INTRODUCTION
Hepatic fibrosis is the result of chronic hepatocellular injury and the first step toward the development of devastating complications such as cirrhosis and hepatocellular carcinoma [1]. The process of hepatic fibrosis is considered to be reversible [2], and the blockade or reversal of hepatic fibrosis has become an important issue in medicine worldwide [3-5]. With advances in knowledge of hepatic fibrosis [6-9], considerable progress has been made in the development of novel drugs; however, most of these drugs are still under clinical trial, with uncertain efficacy and adverse events.
Colostrum is milk from lactating mammals that is secreted during the first approximately 72 hours after birth. Colostrum differs from mature milk in that it contains abundant bioactive components, including growth factors, immunoglobulins, lactoperoxidase, lysozyme, lactoferrin, vitamins, peptides, and oligosaccharides, which are of great importance to human health [10,11]. Thus, colostrum has a variety of beneficial health effects such as bacteriostatic, bactericidal, anti-viral, anti-inflammatory, and immunomodulatory protection [12,13], as well as high antioxidant capacity and radical scavenging activity [14,15].
Oxidative stress is known to be a major contributing factor to the onset of hepatic fibrosis [16]. In pathologic conditions such as chronic hepatitis B virus (HBV) infection, alcoholic liver disease, and non-alcoholic fatty liver disease, the reactive oxygen species (ROS) generated within hepatocytes up- regulate the expression of fibrosis-associated genes, which causes hepatic stellate cell (HSC) activation and collagen deposition. In an experimental setting, carbon tetrachloride (CCl4) is able to induce hepatic fibrosis by stimulating the formation of free radicals, ROSs and lipid peroxidation products [17] and thus is widely used to generate hepatic fibrosis in model animals [18,19].
This study was designed to investigate the preventive effect of colostrum on hepatic fibrosis induced by CCl4 in mice. The study provides a reliable experimental basis for the application of colostrum in the prevention and treatment of hepatic fibrosis.
METHODS
Animals and experimental design
Thirty male (aged 7 weeks) Balb/C mice were purchased from OrientBio (Seongnam, Korea). Following more than 7 days of acclimation, the mice were divided into four groups: the control group (n=5) was fed with water (15 μL/g) five times a week and injected with corn oil (2 μL/g) twice a week intraperitoneally, the bovine colostrum (BC) group (n=5) was fed with BC (15 μL/g) five times a week and injected with corn oil (2 μL/g) twice a week intraperitoneally, the CCl4 group (n=10) was fed with water (15 μL/g) five times a week and injected with CCl4 (1:3 diluted in corn oil, 2 μL/g) twice a week intraperitoneally, and the BC+CCl4 group (n=10) was fed with BC (15 μL/g) five times a week and injected with CCl4 (1:3 diluted in corn oil, 2 μL/g) twice a week intraperitoneally. The mice were treated for 2 weeks and sacrificed 48 hours after the final corn oil or CCl4 injection. When the mice were sacrificed, blood samples and livers were obtained. Blood samples were collected from the inferior vena cava (IVC), centrifuged, and stored at –80°C. All mice were given standard laboratory chow and water ad libitum and maintained under a 12-hour light/dark cycle. All animal experiments were performed according to the guidelines of the Center for Laboratory Animal Research of Samsung Medical Center and with the approval of the Institutional Animal Care and Use Committee of Samsung Medical Center (protocol number: 20140513001).
Materials
BC was purchased from Diatech Korea (Seoul, Korea). CCl4 and corn oil were purchased from Sigma (St. Louis, MO, USA).
Measurements of serum alanine aminotransferase
Alanine aminotransferase (ALT) levels were measured with sera collected from the IVC using an FUJI DRI-CHEM SLIDE GPT/ALP-PIII (Fujifilm Co., Tokyo, Japan) and spectrophotometer (FUJI DRI-CHEM 3000, Fujifilm Co.), according to the manufacturer’s instructions.
Histological examination and immunohistochemistry
Liver samples were fixed in 10% formalin and embedded in paraffin. The sections were then cut (5 μm) and stained with Sirius red to assess fibrosis. For immunohistochemical analysis, the samples were incubated with monoclonal antibody to α-smooth muscle actin (α-SMA) (1:500 dilution, Sigma).
RNA isolation and quantitative polymerase chain reaction analysis
RNA was isolated with a combination of Trizol and subsequent Qiagen RNeasy Plus Mini Kit according to the manufacturer’s instructions (Qiagen, Hilden, Germany). Following reverse transcription, quantitative real-time polymerase chain reaction (qPCR) was performed on an ABI 7300 cycler using TaqMan probe sets for α-actin-2 (Acta2), α-1 type I collagen (Col1α1), tissue inhibitor of metalloproteinase-1 (Timp1), transforming growth factor β1 (Tgfβ1), interleukin 6 (Il6), tumor necrosis factor α (Tnf-α), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Applied Biosystems, Foster City, CA, USA). The expression of each gene was normalized to GAPDH and fold-induction was calculated based on the ΔΔCt method.
Western blot
We performed electrophoresis of protein extracts using T-PER Tissue Protein Extraction Reagent (Thermo Fisher Scientific Inc., Hanover Park, IL, USA) and subsequent blotting. For Western blotting, proteins were transferred to nitrocellulose membranes using an iBLOT blotting system (Life Technologies, Seoul, Korea). We incubated blots with mouse antibody to α-SMA (Sigma) at a dilution of 1:500. Chemiluminscence signals were detected using SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific Inc.). Blots were reprobed with mouse antibody to β-actin (Sigma).
RESULTS
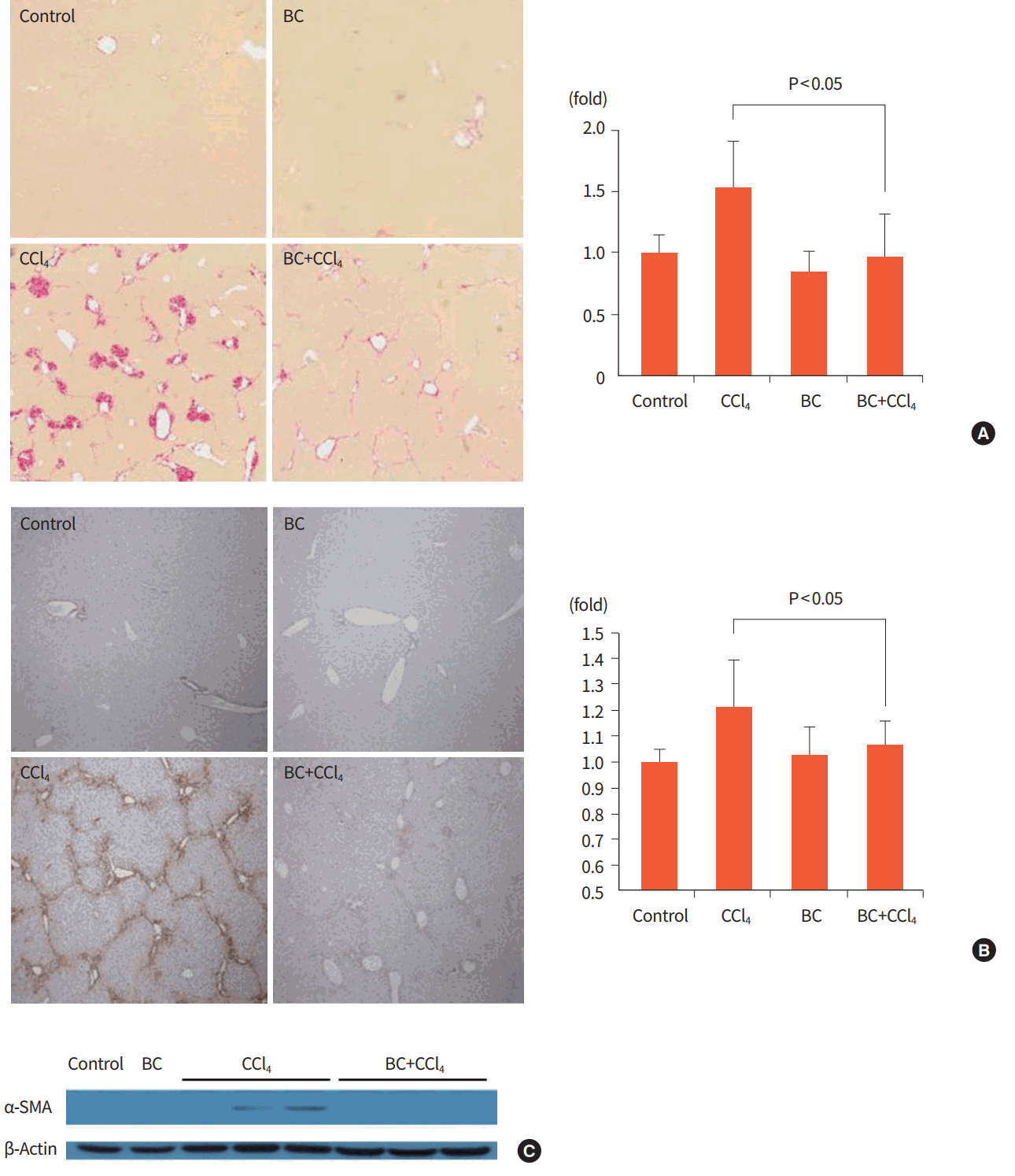
Forty 8 hours after the final CCl4 injection, the CCl4 group showed overt hepatic fibrosis induction by 53% compared to the control group, whereas the BC+CCl4 group had a 55% reduction in fibrosis as demonstrated by Sirius red staining (Fig. 1A).
In the analysis of α-SMA expression, which is a marker of activated HSCs, CCl4 significantly increased α-SMA expression levels compared to those in the control group, whereas additional BC treatment reduced α-SMA expression to the same level as in the control group, as demonstrated by immunohistochemistry and immunoblotting (Fig. 1B, C).
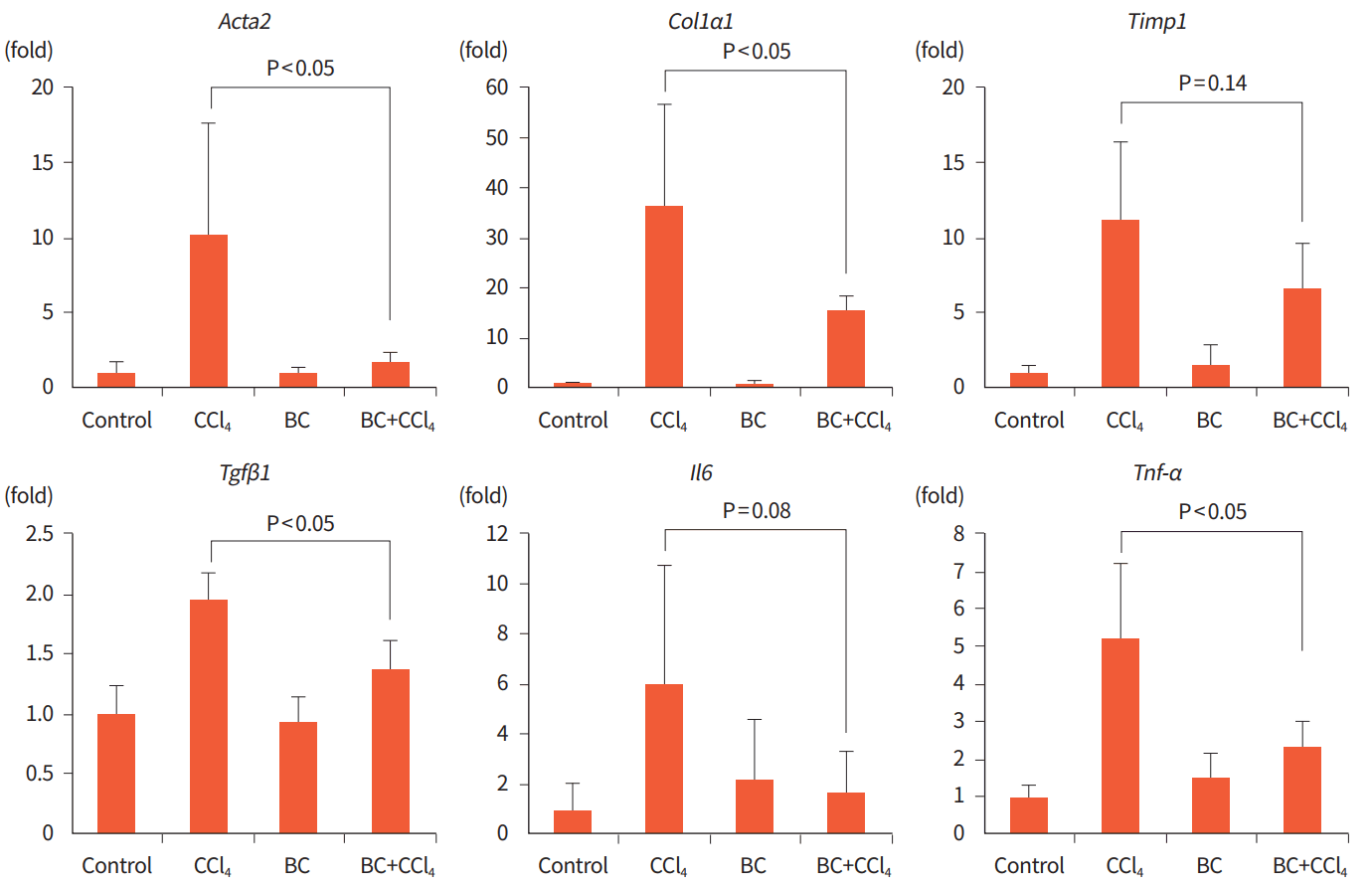
We then analyzed the contribution of BC to the fibrogenic response of the liver. CCl4 treatment increased the expression of Acta2, Col1α1, Timp1, and Tgfβ1, which encode profibrogenic markers, and of proinflammatory cytokine genes Il6 and Tnf-α (Fig. 2). Compared to the CCl4 group, the BC+CCl4 group showed significant 8.42-, 21.11-, 0.57-, and 2.90-fold reductions in the expression of Acta2, Col1α1, Tgfβ1, and Tnf-α, respectively (P<0.05) (Fig. 2). The Timp1 and Il6 expression were reduced by 4.51- and 4.33-fold in the BC+CCl4 group versus the CCl4 group, respectively; however, the differences did not reach statistical significance.
We also found a significant reduction in hepatocellular injury as determined by serum ALT levels in the BC+CCl4 group compared to the CCl4 group (Fig. 3).
DISCUSSION
Oxidative stress is a major factor contributing to the onset of hepatic fibrosis in various liver diseases, including chronic HBV infection and long-term alcohol consumption [16]. ROS are generated mainly via the mitochondrial electron transport chain or via activation of cytochrome P450, mostly cytochrome P450 2E1, nicotinamide adenine dinucleotide phosphate oxidase, or xanthine oxidase or mitochondrial damage. ROS generation in HSC and myofibroblasts occurs in response to several known profibrogenic mediators, including angiotensin II, platelet-derived growth factor, TGFβ, and leptin [20], and the generated ROS up-regulate the expression of critical fibrosis-related genes such as COL1A1, COL1A2, monocyte chemoattractant protein-1 (MCP1), and TIMP1 via activation of signal transduction pathways and transcription factors, including c-Jun N-terminal kinase, activator protein-1, and nuclear factor-κB [1]. Oxidative stress also alters mitochondrial permeability, which leads to the promotion of hepatocyte necrosis and/or apoptosis. Furthermore, hepatocarcinogenesis in the cirrhotic liver is associated with severe oxidative stress [21].
In an experimental setting, CCl4 induces hepatic fibrosis via the formation of free radicals, ROS, and lipid peroxidation products [17]. CCl4 is metabolized into trichloromethyl (–CCl3) and trichloromethyl peroxyl (–OOCCl) by cytochrome P450 enzymes in the liver, and generates free radicals in the process. These free radicals are responsible for oxidative stress- induced hepatic damage and fibrosis [22]. Thus, the CCl4-induced fibrosis model is widely used to evaluate the hepatoprotective effects of various pharmacological therapies. Previous reports have demonstrated that several antioxidant drugs can attenuate CCl4-induced hepatotoxicity [23,24]. The administration of N-acetylcysteine decreases liver fibrosis by increasing glutathione (GSH) level and decreasing the expression of induced nitric oxide synthase (iNOS) [23]. Moreover, treatment with insulin-like growth factor-I reduced myeloperoxidase level and the expression of iNOS in CCl4 model rats [24]. In the present study, we investigated the preventive effect of BC on hepatic fibrosis induced by CCl4. BC dramatically ameliorated CCl4-induced hepatic fibrosis by inhibiting fibrogenesis-related gene expression and also decreased CCl4-induced hepatic injury.
To date, many studies have demonstrated the protective effect of breast milk against various infectious diseases in infants, including respiratory infections, diarrhea, otitis media, and infections in premature infants [25,26]. In addition to the numerous clinically significant benefits of breastfeeding, breastmilk also has bioactive components that protect newborns from hyperoxic challenge due to transition of life to an environment far richer in oxygen than the intrauterine environment [27,28]. It is believed that oxidative stress is involved in the pathogenesis of numerous neonatal diseases such as bronchopulmonary dysplasia, retinopathy of prematurity and necrotizing enterocolitis [29,30]. Breast milk has a number of enzymatic and non-enzymatic antioxidant constituents, including superoxide dismutase, GSH peroxidase, catalase, vitamin E, vitamin C, and β-carotene, which protect newborns against ROS in the early stage of life [29,31]. Accordingly, previous studies have demonstrated the high antioxidant capacity and radical scavenging activity of colostrum. Zarban et al. [14] measured total antioxidant capacity (TAC) and free radical scavenging activity in human milk during the first 6 months of lactation. Both TAC and free radical scavenging activity were significantly higher in colostrum than transitional and mature milk. Quiles et al. [15] also showed that the TAC of milk was significantly higher in colostrum compared with mature and transitional milk. Thus, colostrum might be therapeutically useful for various pathologic conditions in which oxidative stress is involved in the pathogenesis.
In summary, the present study demonstrated that colostrum can ameliorate CCl4-induced hepatic fibrosis by inhibiting fibrogenesis-related gene expression and also decreases CCl4-induced hepatic injury. Therefore, colostrum might be therapeutically efficacious in preventing hepatic fibrosis in a variety of human liver diseases.