INTRODUCTION
Klebsiella pneumoniae is an important pathogen that accounts for one-third of the globally-reported Gram-negative bacterial infections [1]. It causes a wide range of nosocomial infections such as pneumonia, urinary tract infections, bacteremia, and liver abscesses [2]. In particular, antibiotic resistance in K. pneumoniae, including carbapenem resistance, is increasing and has become a critical threat to human health worldwide. The frequency of carbapenem-resistant Enterobacteriaceae, particularly carbapenem-resistant K. pneumoniae which is resistant to carbapenem antibiotics, has significantly risen in the United States and globally over the past decade [3-6]. The lack of novel antibiotics owing to increasing antibiotic resistance has led to the re-evaluation of old antibiotics such as colistin [7].
Colistin is a cationic polypeptide with a lipid tail, mainly targeting anionic lipopolysaccharide (LPS) in the outer membrane of Gram-negative bacteria, causing cell membrane disruption, leakage of cell contents, and cell death [7]. LPS modification with the addition of positively charged residues such as phosphoethanolamine or 4-amino-4-deoxy-1-arabinose is the most common mechanism of colistin resistance in K. pneumoniae [7].
The activation of PhoPQ and PmrAB two-component systems (TCSs) is triggered by environmental signals and specific mutation within the TCSs, which lead to their constitutive activation. As a result, LPS-modifying genes are overexpressed [8]. The PhoPQ indirectly contributes to polymyxin resistance by activating the PmrAB through PmrD, except in certain bacteria like Escherichia coli [9]. The activation of PmrAB leads to the upregulation of pmrCAB and arnBCADTEF-pmrE operons, which facilitate the synthesis and transfer of PetN and L-Ara4N to lipid A [10]. When activated, phosphorylated PmrA binds to the promoter region of the arnBCADTEF operon, improving the recognition of and binding of RNA polymerase and resulting in the upregulation of the operon [11]. In addition to the continuous emergence of colistin resistance, colistin heteroresistance has also been widely reported in K. pneumoniae [12-14].
The heteroresistance phenotype is defined as the subpopulation of bacterial cells with a minimum inhibitory concentration (MIC) level that is higher than the main population’s susceptibility to antibiotics [15]. There are increasing reports that colistin heteroresistance mediates the failure of colistin treatment [16,17]. In addition, it is well-known that antibiotic resistance can be developed from the heteroresistant strain under exposure to antibiotics [18]. Contradictory phenomena are observed in relation to colistin in K. pneumoniae—the first is that the colistin resistance is easily induced from susceptible K. pneumoniae isolates through exposure to colistin [19]. Second, resistant colonies with very high colistin MIC can be selected even by exposure to high concentrations of colistin [13]. This suggests that it is not known whether a colony with very high colistin MIC was selected or a colony with colistin-susceptible to colistin was induced to resistance when exposed to colistin in K. pneumoniae. Previously study also observed that the colistin-resistant population was not well detected in disk diffusion test and E-test in colistin-susceptible K. pneumoniae isolates, but that the colistin-resistant population increased after exposure to colistin [13]. The increase of resistance in colistin-heteroresistant K. pneumoniae isolates may be because of mutation of the susceptible population by stimulation of colistin or owing to the selection of a resistant population by pressure of colistin.
To investigate the origins of the increase in colistin resistance following colistin exposure in colistin-heteroresistant K. pneumoniae, this study generated a colistin-heteroresistant strain artificially, by mixing a colistin-susceptible strain and its resistant mutant. We then tracked the increase or decrease of colistin-resistant and -susceptible populations according to colistin exposure.
This study did not require ethical approvement because it used a reference strain in vitro.
METHODS
Bacterial strains and growth conditions
K. pneumoniae strain American Type Culture Collection (ATCC) 10031 was used for generation of a mimicking colistin-heteroresistant (m-CLHR) strain. The wild-type ATCC 10031 showed a colistin MIC of 0.5 mg/L. The ATCC 10031 with a gentamicin resistance cassette was designated as 10031-CLS in this study. A colistin-resistant mutant (i.e., 10031-CLR) was obtained on media with colistin by increasing the antibiotic concentration serially. The 10031-CLR strain showed a colistin MIC of > 64 mg/L. In addition, this study included 19 colistin-susceptible K. pneumoniae clinical isolates (Table 1); they were obtained from patients in two hospitals of South Korea: the Samsung Changwon Hospital, Changwon and the Samsung Medical Center, Seoul. Bacterial strains were grown in Luria-Bertani (LB) broth (MB cell, BNF Korea) shaking incubated overnight at 37 ℃, 220 rpm and stored in brain heart infusion (BHI) broth (BD, Franklin Lakes, NJ, USA) plus 30% glycerol at –80 ℃ until further analyses. The 10031-CLR strain preserved its colistin MIC by serial culture in an antibiotic-free medium.
Antibiotic susceptibility test
Antibiotic susceptibilities of the K. pneumoniae 10031-CLS, 10031-CLR, m-CLHR, and 19 clinical isolates were determined for colistin and gentamicin using the broth microdilution method as per the Clinical and Laboratory Standards Institute (CLSI) guidelines [20]. The antibiotic breakpoints were determined according to the CLSI breakpoint. E. coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as quality control strains.
Generations of the m-CLHR strain
To differentiate the colistin-susceptible population from the colistin-resistant one, this study used gentamicin resistance as a marker. The gentamicin cassette gene was amplified from pCVD442-gentamicin-resistant (GenR) using primers listed in Table 1. Both polymerase chain reaction (PCR) product and pHK206 were digested with SacI for ligation using Ligation high version 2 (TOYOBO, Kusatsu, Japan), generating a novel plasmid, pJG005. Ligation products were first transformed into chemically competent E. coli DH5α cells. The desired clones were identified by PCR amplification using primers targeting the pHK206 vector backbone (Table 1), and plasmids were extracted for sequencing of the gentamicin cassette gene. The K. pneumoniae ATCC 10031 were streaked on LB agar and incubated overnight at 37 ℃. Electrocompetent cells of K. pneumoniae ATCC 10031 were prepared from a culture incubated in LB at 37 ℃ after the cell density reached an optical density at 600 nm (OD600) of 0.5, and the culture was immediately moved onto ice. Cells were harvested by centrifugation for 20 minutes at 4 ℃ with 4,000 × g and re-suspended in ice-cold sterilized deionized water (DW), followed by washing with ice-cold sterilized DW two times. Finally, cells were re-suspended with icecold sterilized DW. Electrocompetent cells of ATCC 10031 were transformed with purified pJG005 using a Gene Pulser Xcell Microbial System (BIO-RAD) and 1 mm Gene Pulser cuvette (BIO-RAD). The instrument was set at 1.8 kV, 25 µF, 200 Ω, and 1 mm cuvette; transformants of gentamicin-resistant K. pneumoniae ATCC 10031 were screened from the LB plate supplemented with gentamicin 30 mg/L incubated at 37 ℃; the presence of gentamicin resistance cassette was verified by PCR. Thus, the colistin-susceptible ATCC 10031 used in this study, i.e., 10031-CLS, was gentamicin-resistant. In contrast, the colistin-resistant 10031-CLR strain was gentamicin-susceptible.
Survival analysis under high concentration of colistin
After 19 colistin-susceptible K. pneumoniae clinical isolates were exposed to high concentrations of colistin or measured colony-forming unit (CFU)/mL through population analysis profiling (PAP), their survival rates were evaluated. Single colonies of clinical isolates were picked blood agar plates, inoculated into 3 mL of LB broth, and shaking incubated overnight at 37 ℃ with 220 rpm. Overnight cultures were diluted 1:100 into 10 mL of LB broth and sub-cultured at 37 ℃ with 220 rpm until OD600 reached approximately 0.5 (mid-exponential phase) using GeneQuant1300 spectrophotometer (Biochrom). Then, 3 mL of the culture was exposed to 5 mg/L (10-fold MIC) colistin at 37 ℃ with 220 rpm. After colistin treatment for 3 and 9 hours, all samples were washed twice with 1× phosphate-buffered saline (PBS) (Tech & Innovation) and were spotted on antibiotic-free LB agar by serial dilution. The CFU/mL for surviving cell populations was also determined. The experiment was repeated three times independently.
Population analysis profiling
For screening the heteroresistance in 19 colistin-susceptible K. pneumoniae clinical isolates and ATCC 10031 strains including 10031-CLR and m-CLHR, PAPs were performed as previously described [13]. In brief, overnight culture bacterial suspensions were serially diluted to 10-7 with 1× PBS in 96 well plates and plated onto Mueller-Hinton (MH) agar with increasing concentrations of colistin (i.e., 0, 1, 2, 4, 8, 16, and 32 mg/L). After incubation for 18 hours at 37 ℃, strains were classified as heteroresistant if the number of colonies growing at 8-fold that of the MIC was at least 10-7 of those growing on MH agar without colistin [21]. All strains were tested in three times to ensure the accuracy of the entire experiment independently.
Measurement of survival rates after colistin exposure
The survival rate of the colistin-resistant subpopulation was measured before and after exposure to diverse concentrations of colistin [13]. The m-CLHR strain grown in LB broth was inoculated in a 1:100 ratio into MH II broth, with colistin, at 0, 0.125, 0.25, 0.5, 1, 2, 4, 8, 16, and 32 mg/L. After incubating for 24 hours at 37 ℃ with 220 rpm, each bacterial inoculum was diluted with 1 × PBS and spotted on MH agar with and without 4 mg/L of colistin to identify the colistin-resistant cells among the whole population. Colonies were counted after incubation for 18 hours at 37 ℃.
Competition assay
To investigate if the colistin resistance in 10031-CLR and the plasmid with gentamicin resistance cassette in 10031-CLS confers fitness cost, we performed a competition assay based on two ratios of 10031-CLS and 10031-CLR, 1:1 and 1:10-7. For the competition assay, single colonies of 10031-CLS and 10031-CLR were picked from LB agar, inoculated into 3 mL of LB broth, and shaking incubated overnight at 37 ℃ with 220 rpm. All cultures were then back-diluted in LB to an OD600 of 2.0 (approximately 109 CFU/mL) to normalize cell density across mix conditions in 1:1 and 1:10-7 ratios. After that, the mixtures were inoculated in a 1:100 ratio into fresh LB broth and were shaking incubated for 18 hours at 37 ℃ with 220 rpm. Cultures were diluted with 1× PBS (Tech & Innovation) and spotted on MH agar with and without 4 mg/L of colistin. The colonies surviving in MH agar were counted after 18 hours of culture. The competition index was then defined as the ratio of CFU/mL of the resistant cells to CFU/mL of the susceptible cells. All experiments were repeated three times independently.
Fluctuation and stability tests for the m-CLHR strain
For the fluctuation test, the m-CLHR strain was inoculated in 1:100 ratio in 10 mL of eight LB broth tubes, and was shaking incubated overnight at 37 ℃ with 220 rpm on a shaking incubator. Each overnight culture was spotted onto MH agar with and without 4 mg/L of colistin. The colonies surviving in MH agar were counted after 18 hours of culture. The ratio of CFU/mL of resistant cells to the CFU/mL of susceptible cells was calculated for each of the eight tubes.
Following the fluctuation test, the stability of resistant populations in m-CLHR strain was investigated. Cultures of eight tubes in the fluctuation test were repeatedly sub-cultured for additional four days in the absence of colistin. The ratio of resistant cells to the susceptible cells were measured for each serial subculture.
In vitro time-killing assay
In vitro time-killing assay for colistin was performed for the m-CLHR strain. The cultured strain was inoculated in a 1:100 ratio into 10 mL of freshly prepared LB broth supplemented with 4 mL colistin and shaking incubated at 37 ℃ with 220 rpm. Following this, 200 µL aliquots of each bacterial inoculum were collected at 0, 1, 2, 4, 8, 12, and 24 hours and were diluted with 1 × PBS. Bacterial cultures were spotted on three different MH agars: those with no antibiotics, with 30 mg/L gentamicin, and with 4 mg/L colistin to differentiate the 10031-CLS and 10031-CLR, respectively. All experiments were repeated three times independently.
Sequence analyses
We determined nucleotides and deduced amino acid alteration in genes associated with colistin resistance. PCR and DNA sequencing were performed for phoPQ, pmrAB, mgrB, and pbgP. These have been known to be associated with colistin resistance in 10031-CLS, 10031-CLR, and the colistin-resistant colony obtained in in vitro time-killing assay [19].
RESULTS
Colistin heteroresistance in K. pneumoniae clinical isolates
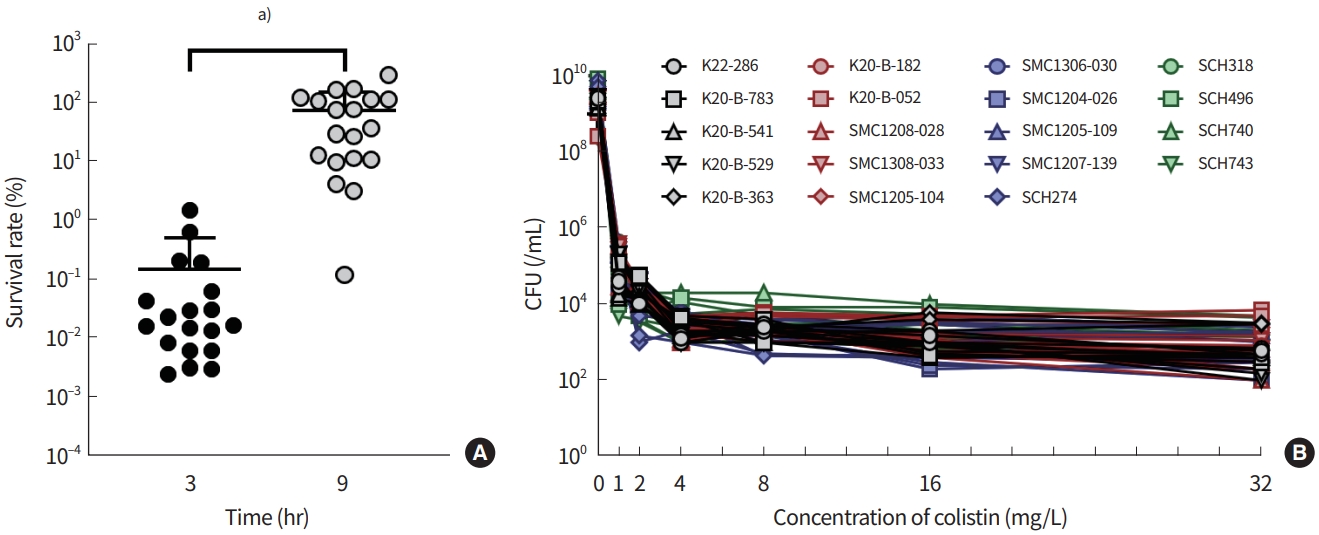
All K. pneumoniae clinical isolates and reference strain ATCC 10031 were susceptible to colistin (Table 2). After being exposed to a 5 mg/L (10-fold MIC) of colistin, the clinical isolates showed a decrease in survival rates within 3 hours. However, all of the isolates exhibited regrowth after 9 hours of exposure to colistin (Fig. 1A). PAP showed that all the K. pneumoniae clinical isolates were colistin-heteroresistant; that is, subpopulations survived at a frequency of > 10-7 in the MH agar with eight times the colistin MICs (Fig. 1B).
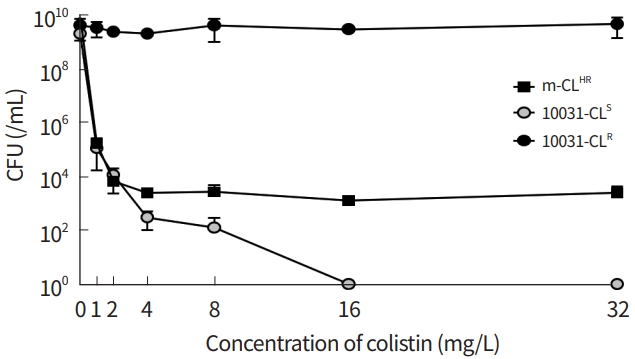
A colistin-susceptible and gentamicin-resistant strain, i.e., 10031-CLS, was obtained from the K. pneumoniae reference strain ATCC 10031 by transforming a plasmid with gentamicin resistance cassette. By mixing the 10031-CLS and 10031-CLR at a ratio of 1:100, a m-CLHR strain was generated (Table 1) [22]. We performed PAP for 10031-CLS, 10031-CLR, and m-CLHR, respectively, and the m-CLHR showed the heteroresistance phenotype (Fig. 2).
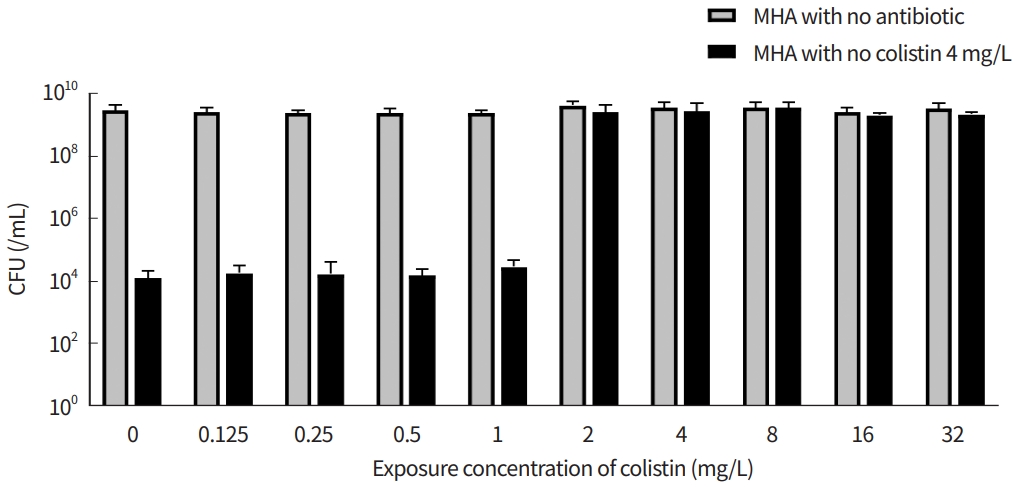
The survival rates of colistin-resistant cells in m-CLHR were investigated both before and after colistin exposure (Fig. 3). When the m-CLHR was treated with 1 mg/L colistin, the survival rates of the resistant cells were preserved. However, the proportion of resistant cells among the total population reached almost 100% when colistin was treated with more than 2 mg/L. Although this pattern is typical heteroresistance [13,23], it is unclear whether a resistant population is selected or whether a susceptible population has become resistant due to mutation.
Maintenance of heteroresistance in m-CLHR
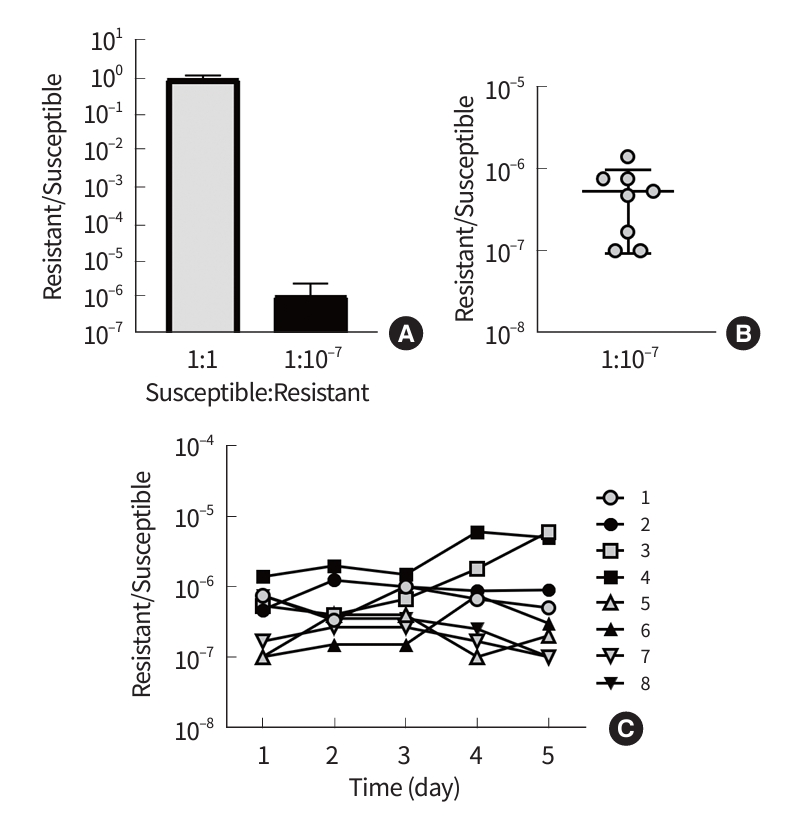
During the competition assay, neither 10031-CLS nor 10031-CLR showed any fitness cost (Fig. 4A). When they were mixed in a 1:1 ratio, the ratio of survived resistant cells to survived susceptible cells was about 1, indicating no fitness cost for both. The ratio of resistant cells increased to about 10-6 when they were mixed with 1:10-7; however, its increase was estimated to be not statistically significant.
In addition, the ratio of resistant cells to susceptible cells, i.e., 1.0× 10-7 to 1.4× 10-6 (average, 5.3× 10-6), did not deviate significantly from the range of heteroresistance in the fluctuation test (Fig. 4B). All eight lineages obtained in the fluctuation test on m-CLHR were subjected to investigation for stability of resistant cell ratio. Although one lineage (i.e., lineage #3) showed an increasing tendency of resistant cells, the ratios of resistant cells preserved their stability in most lineages despite several days of subculture in antibiotic-free media (Fig. 4C).
In vitro time-killing assay for m-CLHR
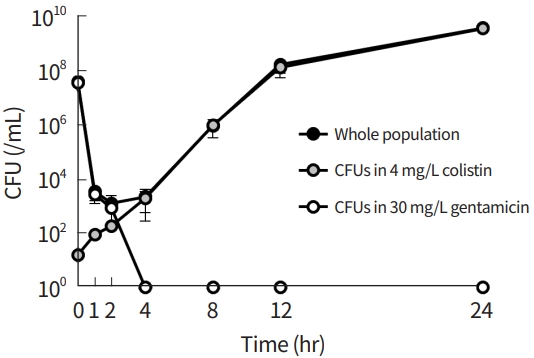
An in vitro time-killing assay was performed for m-CLHR, and changes in 10031-CLS and 10031-CLR populations within the m-CRHR were measured over time using different selection media (Fig. 5). While the colistin-susceptible cells, i.e., 10031-CLS, were selected in the media with 30 mg/L gentamicin, the resistant cells, i.e., 10031-CLR, were found in the media with 4 mg/L colistin. Prior to colistin treatment, the ratio of resistant cells to susceptible cells was estimated to be about 10-7, corresponding to the original proportion of the mixture. The number of total bacterial cells decreased initially by treatment of 4 mg/L colistin, but regrowth typical to heteroresistance was observed after 4 hours upon exposure to colistin. CFUs in the media with 30 mg/L decreased with time, and all susceptible cells were eliminated after 4 hours of colistin treatment. In contrast, CFUs in the media with 4 mg/L increased with time, and resistant cells accounted for the entire surviving population.
The surviving colony in the in vitro time-killing assay showed the same nucleotide substitution, i.e., G1265→T1265 in phoQ resulting in Ser422→Ile422.
DISCUSSION
This study was performed because two incongruent phenomena on colistin resistance in K. pneumoniae were observed. One is that colistin resistance is very easily induced from susceptible K. pneumoniae through colistin exposure, irrespective of its concentrations [19]. The other is that colistin-heteroresistant isolates are not well identified based on classical methods [17,24]. These phenomena raise the question of whether colistin resistance occurs from colistin-heteroresistant K. pneumoniae strains after exposure to colistin. When a colistin-heteroresistant strain is treated with colistin, the re-grown population after a certain period of time may have developed resistance through mutation or overexpression in colistin associated genes caused by colistin stimulation, or only colistin-resistant subpopulation that already existed in the colistin-heteroresistant strains may have survived.
This study modeled a colistin-heteroresistant K. pneumoniae strain to investigate the origin of surviving resistant population posterior to colistin treatment. The modeled heteroresistant strain was generated by mixing a colistin-susceptible strain and a colistin-resistant mutant obtained from the susceptible strain. To differentiate between the two populations, this study introduced gentamicin resistance cassette into the colistin-susceptible strain. The two populations showed no differences in survival fitness on media without antibiotics. In addition, the modeled colistin-heteroresistant K. pneumoniae strain showed a typical heteroresistance pattern in PAP, and maintained its heteroresistance on antibiotic-free media.
As a result, this study identified only the colistin-resistant population that survived after antibiotic treatment in the modeled colistin-heteroresistant K. pneumoniae strain. Results of this research indicate that the development of colistin resistance from a heteroresistant strain is likely because of the selection of the resistant population. Previous studies on the heteroresistance and resistance presumably suggest that resistant populations survive among the heteroresistant strains with no experimental basis. Our study presents the experimental evidence on the emergence of resistance from heteroresistant strain. Thus, the rapid emergence of colistin resistance from colistin-susceptible K. pneumoniae would be the result of overgrowth of the resistant population that already exists in the susceptible strain.
Previous research has indicated that colistin heteroresistance may be prevalent in K. pneumoniae, although it could not be detected easily [13,17,24]. The presence of resistant populations in phenotypically colistin-susceptible K. pneumoniae isolates seems to be an intrinsic feature, and colistin treatment would trigger both colistin resistance and treatment failure [18,25].
Although this study investigated only one K. pneumoniae strain, it can provide meaningful experimental evidence for the mechanism through which colistin resistance emerges or increases after exposure to it.